Vasi malpighiani. Il significato dei vasi malpighiani nell'enciclopedia della biologia. Struttura dei vasi malpighiani
Nelle zecche ixodidi, le funzioni di rimozione dell'acqua e dei sali in eccesso dal corpo sono eseguite dalle ghiandole salivari e i prodotti del metabolismo dell'azoto sono eseguiti dai vasi malpighiani. I vasi malpighiani sono escrescenze tubolari dell'intestino medio. Si estendono dalla parte anteroventrale della vescica rettale ai lati del retto, tornano indietro e raggiungono la parte posteriore dell'idiosoma (Fig. 9). Poi si rivolgono in avanti e verso l'alto e lungo il lato dorsale degli organi interni raggiungono la base dello gnatosoma, tornano indietro e lungo il lato ventrale sotto i processi intestinali si dirigono medialmente dalle ghiandole salivari al livello del singanglio.
Per tutta la loro lunghezza, ad eccezione delle estremità distali leggermente espanse ("fiale"), sembrano tubi bianchi o trasparenti quasi uniformi e possono variare nel grado di carico di escrementi delle singole aree. All'esterno i vasi sono circondati da una membrana di tessuto connettivo e da un plesso di fibre muscolari. Le loro pareti per tutta la loro lunghezza sono formate da un unico strato di cellule epiteliali, che appaiono omogenee alle colorazioni istologiche convenzionali. Con la colorazione istochimica dei mucopolisaccaridi nella zecca della taiga, è stato possibile distinguere due tipi di cellule epiteliali nelle parti distali, alcune delle quali contenevano inclusioni di mucopolisaccaridi neutri e altre di mucopolisaccaridi acidi. Entrambi i tipi di cellule si presentano senza alcuno schema. La cavità del vaso contiene cristalli escretori sferici (Tav. XXIII, 7), che rappresentano apparentemente la guanina e, forse, un'altra purina non identificata, come è stato stabilito per diverse specie della sottofamiglia. Amblyomminae (Hamdy, 1973).
Durante l'esame al microscopio elettronico (Atlas..., 1979), le cellule epiteliali delle pareti vascolari mostrano la polarità caratteristica degli organi emuntori. Le loro superfici apicali, rivolte verso il lume del vaso, recano microvilli e la membrana citoplasmatica basale forma numerose invaginazioni (“labirinto”) (Tabella XXIII, 2). In base al riempimento del citoplasma con inclusioni, al grado di sviluppo dei microvilli, al labirinto basale e ad alcune altre caratteristiche della nave malpighiana, si distinguono 3 sezioni.
Le parti distali dei vasi sono formate da cellule, il cui citoplasma, soprattutto nella zona apicale, contiene un gran numero di corpi delimitati da membrana pieni di una sostanza a grana fine di natura mucopolisaccaridica. Molte cellule delle sezioni distali, e soprattutto le loro estensioni terminali a forma di ampolla, contengono molte rickettsie simbiotiche del genere Wolbachia (Tabella XL). Le cellule della sezione principale, successiva a quella distale, sono caratterizzate da un moderato sviluppo dei microvilli e del labirinto basale, nonché dall'assenza di inclusioni mucopolisaccaridiche nel citoplasma. Allo stesso tempo, queste cellule contengono un gran numero di granuli di glicogeno e inclusioni grasse. La cavità dei vasi contiene un gran numero di sferiti escretori arrotondati con stratificazione ben definita (Tav. XXIII, 3). Le sezioni prossimali sono caratterizzate dal più intenso accumulo di sferiti di guanina nella loro cavità, circondata da cellule con numerosi e lunghi microvilli.
La vescica rettale è una sacca spaiata situata alla giunzione dell'intestino medio con l'intestino posteriore. La sua forma e dimensione dipendono in gran parte dal grado di riempimento della cavità vescicale con feci e prodotti di escrezione dei vasi malpighiani. Si presume che nella vescica rettale, come nelle parti prossimali dei vasi, avvenga il riassorbimento dell'acqua dagli escrementi e dalle feci.
Il prodotto più importante del metabolismo dell'azoto, l'ixoduro - guanina, è caratterizzato da una solubilità estremamente bassa. Si presume che nelle cellule della sezione distale sia rappresentato dal suo precursore più solubile: la guanosina. Nella cavità delle parti principali e prossimali dei vasi, la guanina viene scissa dalla guanosina, che forma caratteristici cristalli sferici. Inoltre, nelle stesse sezioni prossimali, così come nella vescica rettale, avviene il riassorbimento dei principali componenti del fluido escretore, per cui il contenuto della vescica è rappresentato da una densa massa viscosa di guanina ed, eventualmente, altri prodotti del metabolismo dell’azoto.
Il grado di riempimento dei vasi malpighiani e della vescica rettale con la guanina varia notevolmente a seconda dello stato fisiologico della zecca. Ad esempio, nelle femmine affamate di 1. persulcatus (Taiga tick..., 1985), recentemente mutate, le pareti dei vasi sono allungate con contenuto bianco brillante (guanina) e hanno un diametro di 50-70 micron. Lo scarico degli escrementi accumulati durante il periodo di preparazione alla muta avviene durante la prima settimana dopo la muta. Successivamente i vasi appaiono traslucidi, hanno un diametro di 40-60 micron e contengono solo accumuli locali di sferiti escretori. Il numero di questi ultimi negli individui affamati a lungo termine aumenta notevolmente, tanto che nei vasi si alternano aree trasparenti “vuote” e aree bianche piene di escrementi. Durante l'alimentazione delle femmine, durante i primi 2 giorni, i vasi vengono ripuliti dagli escrementi, ma poi iniziano rapidamente a riempirsi nuovamente di guanina. Nelle zecche nutrite, il diametro dei vasi sanguigni è 3-4 volte maggiore rispetto alle zecche affamate. Dopo che le femmine si allontanano dall'ospite, il caricamento dei vasi con escrementi avviene in modo ancora più intenso e il loro diametro può aumentare 10 o più volte rispetto agli individui affamati e raggiungere i 500-800 micron. Sono riempiti per tutta la loro lunghezza da una massa bianca di guanina. Anche la vescica rettale delle femmine ingorgate aumenta notevolmente, la defecazione si interrompe e la guanina rimane nel corpo fino alla morte della zecca.
Nelle larve e nelle ninfe che sono in congestione e si preparano alla muta, a differenza delle femmine, i vasi malpighiani appaiono trasparenti, poiché il loro contenuto viene immediatamente spinto nella vescica rettale. In preparazione alla muta del rivestimento cuticolare del retto, la comunicazione della vescica rettale con l'ambiente esterno viene interrotta e prima della fine della muta la defecazione diventa impossibile. Di conseguenza, alla fine della muta, la vescica rettale è piena di guanina e occupa la maggior parte della metà posteriore del corpo.
La defecazione della zecca della taiga avviene con una certa ciclicità temporale e allo stesso tempo cambia la composizione delle feci. Nelle zecche affamate, la defecazione è più intensa nei primi giorni dopo la muta e le feci sono di colore bianco brillante e apparentemente sono costituite quasi solo da guanina. Successivamente, nelle zecche affamate, diventano bianche, marroni o nere, a seconda del contenuto di ematina, proveniente dall'intestino medio. La defecazione è più intensa durante l'alimentazione. La materia fecale in questo momento è marrone, nera o rosso scuro a causa dell'escrezione di grandi quantità di emoglobina e sangue intero.
(insetti e millepiedi) e aracnidi. Apparentemente, in questi gruppi sono sorti indipendentemente: dall'ectoderma negli insetti e nei millepiedi e dall'endoderma negli aracnidi. A volte le escrescenze intestinali trovate nei tardigradi vicino agli artropodi sono considerate vasi malpighiani (Dogel, 1981). Prendono il nome dal biologo Malpighi, che li scoprì nel 1669.
Struttura
I vasi malpighiani sono tubuli epiteliali chiusi ciecamente che si aprono al confine dell'intestino medio e posteriore. Tuttavia, in alcuni (Nicrophorus, Gnaptor, Coccoidea) si aprono nell'intestino medio, in altri (larve di farfalle, coleotteri di bronzo, formiche di leone) - nell'intestino posteriore.
Il numero dei vasi malpighiani varia da 2 a 150 (nell'ape), sono poco sviluppati nelle proture e completamente assenti nei collemboli, negli afidi sono secondariamente ridotti;
I vasi malpighiani possono terminare liberamente nell'emolinfa, in alcuni casi crescono insieme a coppie, formando anse chiuse; A volte le punte dei tubuli perforano il rivestimento muscolare dell'intestino posteriore. Questo fenomeno si chiama criptonefria(negli scarabei, merletti, bruchi e larve di seghe). La fusione dei vasi malpighiani con gli intestini si osserva per i rappresentanti terrestri costretti alla conservazione dell'acqua. La parte criptonefrica del tubulo escretore aiuta ad attingere acqua dall'intestino posteriore. Negli insetti che vivono nell'acqua o nel suolo, non si osserva la criptonefria.
Le pareti dei vasi sanguigni sono costituite da epitelio a strato singolo. Le celle sono sparse, soprattutto nella sezione terminale. Come l'epitelio dell'intestino medio, la superficie delle cellule rivolta verso il lume reca il rabdorio, il suo strato di bastoncini plasmatici. All'esterno l'epitelio è ricoperto da una membrana basale; i vasi sono dotati di muscoli propri e sono in grado di piegarsi.
Ogni nave è funzionalmente divisa in due parti. La sezione terminale consente l'ingresso dell'acqua e dei sali solubili dell'acido urico provenienti dall'emolinfa. Successivamente si spostano alla base della nave, in un ambiente ricco di CO 2. L'acido urico viene spostato dal sale carbonico e precipita, mentre il bicarbonato entra nell'emolinfa. I cristalli di acido urico si spostano verso l'uscita dell'intestino e vengono poi eliminati con le feci. Questo funzionamento del vaso malpighiano è stato descritto da Shvanvich per l'insetto Rhodnius, mentre qualcos'altro è possibile: quando si verifica la perdita di cristalli di acido urico all'interno delle cellule del vaso, e il canale effettua solo l'uscita.
Funzioni
escretore
I vasi malpighiani di insetti e millepiedi liberano il corpo dall'azoto in eccesso principalmente sotto forma di acido urico e negli aracnidi sotto forma di guanina (Dogel, 1981). Entrambe le sostanze sono eterocicli di due anelli, composti con un rapporto azoto/idrogeno maggiore di quello dell'urea (un prodotto dell'escrezione dei mammiferi). Pertanto, è più adatto per il risparmio idrico.
Oltre all'acido urico, i vasi malpighiani sono in grado di secernere ione ammonio, urea e acido salicilico (rilevante per gli insetti che si nutrono di Salix).
Le funzioni escretorie dei vasi malpighiani sono strettamente correlate alle funzioni dell'intestino posteriore. Nel caso più semplice, i vasi malpighiani aspirano solo il plasma emolinfatico e lo trasferiscono nell'intestino posteriore. Tutti gli altri compiti sono svolti dalle papille rettali dell'intestino posteriore, che restituiscono acqua e altre sostanze utili all'emolinfa e rimuovono gli escrementi disidratati e le molecole "extra" dal corpo. In molti insetti i vasi malpighiani partecipano anche all'aspirazione inversa dell'acqua, formando un unico complesso di organi emuntori con l'intestino posteriore.
Altre funzioni dei vasi malpighiani
Oltre alla secrezione, i vasi malpighiani possono svolgere altre funzioni. In un certo numero di insetti (ortotteri, coleotteri macinati, coleotteri carnivori), si può formare l'enzima digestivo dipeptidasi (Shvanvich, 1949).
I vasi malpighiani di alcuni insetti possono secernere granuli di carbonato di calcio. Nelle mosche si scioglie nel sangue durante la pupa e si deposita poi nelle pareti del pupario. Una dissoluzione simile avviene negli insetti stecco, ma il calcio entra poi nella parete del corion dell'uovo.
Nelle larve di formicaleone e crisopo, prima della pupa, i vasi malpighiani secernono seta, dalla quale la larva tesse il suo bozzolo. Al momento della secrezione della seta, le cellule diventano multinucleate.
Lo stesso acido urico escretore può avere anche funzioni aggiuntive. Nei bianchi, nei merletti, nelle vespe e nei sirfidi, viene utilizzato per costruire pigmenti di natura purina: bianco, giallo e arancione. Nelle larve di pennitsa, l'acido urico, insieme all'enzima saponificante, viene escreto negli escrementi e si combina con la cera secreta da una ghiandola speciale, formando una schiuma protettiva.
Riferimenti
- Tyshchenko V.P. Fisiologia degli insetti. M.: Scuola superiore, 1986. 303 p.
- Shvanvich B.N. Corso di entomologia generale. M.-L.: Scienza sovietica, 1949. 895 p.
- Dogel V.A. Zoologia degli invertebrati. 7a edizione, rivista. e aggiuntivi M.: Scuola superiore, 1981. 606 p.
- Kluge N.Yu. Tassonomia moderna degli insetti. Principi di tassonomia degli organismi viventi e sistema generale degli insetti con classificazione degli insetti alati primari e antichi. San Pietroburgo: Lan, 2000. 336 p.
Dizionario dell'apicoltore
Vasi malpighiani
Gli organi dell'apparato escretore dell'ape, situati nella cavità addominale, hanno l'aspetto di lunghi tubi sottili, 100-150 nelle api adulte, 4 nelle larve, le secrezioni dei vasi malpighiani vengono utilizzate dalla larva per tessere un bozzolo.
Enciclopedia "Biologia"
Vasi malpighiani
Escrescenze tubolari cieche, a volte ramificate, dell'intestino nei ragni e negli insetti. Svolge le funzioni di escrezione e osmoregolazione. I ragni hanno 1-2 paia di vasi malpighiani, che sono sporgenze dell'intestino medio vicino alla cloaca. Negli insetti sono più numerosi (fino a 150 coppie) e si formano al confine dell'intestino medio e posteriore. Le pareti dei vasi malpighiani assorbono i prodotti metabolici liquidi dalla cavità corporea nei lumi di questi vasi si formano piccoli cristalli di acido urico, che vengono escreti nell'intestino posteriore; Gli insetti che vivono in ambienti acquatici e umidi, così come quelli che si nutrono di cibi vegetali succulenti, possiedono il maggior numero di vasi malpighiani. Rimuovono l'acqua in eccesso dal corpo, cioè sono osmoregolatori.
Dizionario enciclopedico
Vasi malpighiani
(denominato M. Malpighi), organi emuntori di aracnidi, millepiedi e insetti. Lunghe escrescenze tubolari dell'intestino al confine dell'intestino medio e posteriore. Negli insetti acquatici partecipano all'osmoregolazione.
I vasi malpighiani prendono il nome dal naturalista italiano M. Malpighi, che li scoprì negli insetti nel XVII secolo. A proposito, questo stesso scienziato è noto anche in relazione ai risultati ottenuti nell'anatomia e nella fisiologia umana: nel 1661. dimostrò che il sangue tra le vene e le arterie passa attraverso i capillari e scoprì la microcircolazione.
Struttura dei vasi malpighiani
I vasi malpighiani si presentano sotto forma di tubi, che da un lato terminano ciecamente nel corpo e dall'altro sfociano nel canale intestinale, solitamente tra l'intestino medio e quello posteriore. (foto)
In alcuni insetti, il luogo della loro confluenza è leggermente spostato rispetto a quello tipico. Ad esempio, negli scarabei Necrophorus, i vasi si aprono nella cavità dell'intestino medio davanti alla valvola pilorica (la formazione che la separa dall'intestino posteriore). Nell'antleone, i bruchi e gli scarabei di bronzo, al contrario, confluiscono nell'intestino posteriore. Infine, lo scarabeo fogliare è diverso da tutti gli altri: ha 6 vasi, i primi due confluiscono nell'intestino medio e i restanti 4 comunicano con l'intestino posteriore.
I tubi si aprono nell'intestino separatamente o sono collegati in fasci con dotti comuni. Nelle libellule, i vasi vengono raccolti in 4 fasci e, ad esempio, negli scarafaggi - in 6.
Le parti libere dei vasi che terminano nel corpo sono sospese e ne aspirano gli escrementi nocivi. Nei bruchi e in numerosi coleotteri, le estremità dei tubi sono attaccate all'intestino posteriore, assorbono l'acqua da esso, migliorando il lavoro delle ghiandole rettali, che di solito svolgono questa funzione. Questo fenomeno è chiamato criptonefria.
Il diametro dei vasi per tutta la loro lunghezza è solitamente lo stesso, ma a volte possono esserci espansioni (serbatoi) in parti diverse di essi. In alcuni gruppi i tubi si presentano sotto forma di rami: irregolari (ad albero) o piumati.
Il numero totale di tubuli malpighiani negli insetti varia ampiamente. Afidi e Collemboli non li hanno affatto; Gli insetti ne hanno due, i Tripidi e la maggior parte delle Mosche ne hanno due paia, la maggior parte dei Lepidotteri ne ha 6. Esistono anche possessori di un maggior numero di organi emuntori: le Libellule hanno 20 vasi malpighiani, nelle Blatte il loro numero arriva a 90-120, nelle api fino a 150. La locusta del deserto ha il maggior numero di tubi: 250.
L'origine esatta dei vasi malpighiani è sconosciuta e gli esperti non hanno un parere definitivo al riguardo.
Funzioni dei vasi malpighiani
Questi sono gli organi escretori; pertanto, la loro funzione principale è quella di rimuovere dal corpo degli insetti i metaboliti che non possono essere depositati o inclusi in altri processi biochimici. Principalmente, i vasi rimuovono l'acido urico dal corpo. Questo processo può essere rappresentato come segue.
Oltre all'acido urico, tra gli escrementi si trovano altri composti, come l'aminoacido leucina o il carbonato di calcio. La cicalina australiana (Ptyelis) costruisce addirittura un guscio a spirale calcareo dalle secrezioni dei vasi malpighiani contenenti molto calcio. Calliphora e lucilia, essendo allo stadio larvale, rilasciano ammoniaca in forma libera. Inoltre, gli escrementi di molti insetti contengono sali di ammonio, allantoina (nei moscerini), creatina (in Rhodnius) e acido salicilico (negli insetti che vivono sul salice o sul pioppo, la cui linfa contiene questo composto).
Oltre all'escrezione, sono possibili ulteriori funzioni dei vasi malpighiani. In alcuni secernono enzimi digestivi, ad esempio negli scarabei macinati secernono dipeptidasi. Ulteriori “porzioni” di questo enzima, che scompone le proteine, sono necessarie perché questo insetto predatore si nutre principalmente di alimenti animali a base di proteine. (foto)
Nelle larve dell'Antleone i vasi malpighiani producono seta; in altre, una sostanza calcarea (il bozzolo ne resta impregnato quando si trasforma in pupa). Nel punteruolo Phytonomus, i vasi malpighiani secernono seta anche dalle loro sezioni centrali, dalla quale tessono un bozzolo, e nella sezione principale si forma un composto con il quale questo bozzolo viene lavorato dall'interno.
Per molti organi escretori di invertebrati non è ancora stata risolta la questione se l'urina primaria sia formata mediante ultrafiltrazione. Ma per gli organi emuntori degli insetti la risposta sembra essere decisamente negativa; Si basa sul fatto che l'inulina, considerata l'indicatore più affidabile dell'ultrafiltrazione, non passa nell'urina degli insetti, il che indica l'assenza di ultrafiltrazione (Ramsay e Riegel, 1961). Come si forma allora l'urina negli insetti?
Il loro sistema escretore è costituito da tubi chiamati vasi malpighiani, che possono essere da due a diverse centinaia. Ciascun vaso si apre nell'intestino al confine tra l'intestino medio e l'intestino posteriore; l'altra estremità è cieca e nella maggior parte degli insetti si trova nell'emocele (Fig. 10.6). Ma alcuni insetti, in particolare gli scarabei che si nutrono di sostanza secca (ad esempio le larve del verme della farina Tenebrio), hanno un adattamento speciale che fornisce una straordinaria capacità di estrarre l'acqua dagli escrementi. In essi, l'estremità cieca del vaso malpighiano è strettamente collegata al retto e l'intera struttura è circondata da una membrana (membrana perirettale). Lo spazio che forma è pieno di fluido perirettale, che circonda sia il vaso malpighiano che l'epitelio rettale, ma è separato dall'emolinfa generale.
La nave malpighiana funziona come segue. Il potassio viene trasportato attivamente nel lume della nave, seguito passivamente dall'acqua sotto l'influenza delle forze osmotiche. Di conseguenza, nel lume si accumula molto liquido ricco di potassio e da qui passa nell'intestino posteriore. Ci sono le gare
le sostanze disciolte e una parte significativa dell'acqua vengono riassorbite e l'acido urico (entrato nel liquido sotto forma di urato di potassio idrosolubile) precipita. Ciò facilita l'ulteriore estrazione dell'acqua, poiché l'acido urico precipitato non partecipa all'attività osmotica del contenuto intestinale. E cosa succede? rimane nel retto* e alla fine viene escreto come una miscela di urina e feci.
Gli insetti che si nutrono di vegetazione fresca mangiano cibo ad alto contenuto di acqua ed espellono molta urina liquida. E coloro che consumano cibo secco sono in grado di espellere escrementi molto secchi, quindi difficilmente perdono acqua nelle urine e nelle feci. Un buon esempio è la larva della cocciniglia (Tenebrio moli~ tor). L'intero ciclo di vita di questo insetto
Riso. 10.6. Sistema escretore della cimice Rhodnius. (Wiggles-worth, 1931.)
Solo il ®dkn* dei quattro vasi malpighiani è riportato per intero. Queste navi sono divise in due parti diverse: superiore e inferiore. Si svuotano nel retto alla sua giunzione con l'intestino medio.
passa nella farina secca, eppure non gli manca l'acqua.
La larva del verme della farina ha la tipica membrana perirettale sopra descritta e gli escrementi prodotti possono essere così secchi da assorbire acqua dall'aria con un'umidità relativa del 90% (Ramsay, 1964). Vediamo come. Riesci a immaginare il meccanismo per estrarre l'acqua? La concentrazione osmotica del fluido nello spazio perirettale può
può essere molto elevato [la depressione del punto di congelamento (FDP) raggiunge gli 8°] - molto più alta della concentrazione dell'emolinfa (dove l'AT varia da 0,7 a 1,4°). La differenza è più marcata quando il verme della farina viene tenuto in un ambiente con un’umidità molto bassa.
Esistono due possibili modi di funzionamento del complesso rettale nella larva del verme della farina: 1) trasporto attivo dell'acqua 
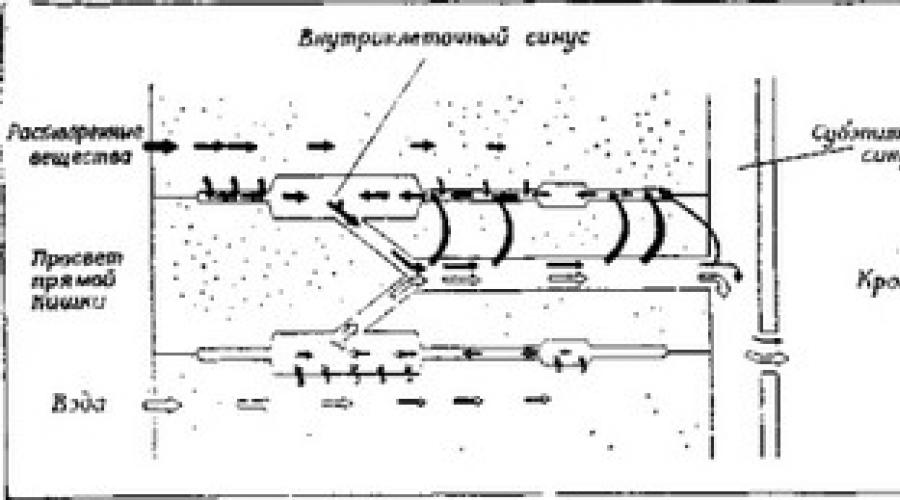
Riso. 10.7. Modello del movimento dell'acqua nella bistecca rettale dello scarafaggio Periplaneta americana. (Oschman, Muro, 1969.)
Il movimento delle sostanze disciolte è indicato dalle frecce nere, il movimento dell'acqua dalle frecce chiare. Questi movimenti sono presentati separatamente solo per chiarezza, ma in realtà si verificano nello stesso seno intracellulare. Le sostanze disciolte vengono pompate attivamente nel seno intracellulare sia dal lume intestinale che dal fluido che scorre nell'eino subepiteliale. L'elevata concentrazione osmotica nel seno intracellulare provoca un afflusso osmotico di acqua, che porta al passaggio di acqua e sali nel seno sottoepiteliale. Questo sistema è simile al modello Curran a tre compartimenti (Figura 9.14).
dal lume di questo complesso nell'emolinfa o 2) trasporto attivo di alcuni soluti (presumibilmente cloruro di potassio) dall'emolinfa nello spazio perirettale, aumentando la concentrazione del fluido perirettale e quindi portando alla rimozione osmotica dell'acqua dal retto (Grimstone et al., 1968).
Il secondo di questi meccanismi di riassorbimento dell'acqua è identico in linea di principio a quello postulato nella teoria dei tre compartimenti di Curran (Capitolo 9). Studiando gli scarafaggi, sono state ottenute ulteriori prove a favore di questa ipotesi. Se lo scarafaggio è in grado di bere acqua, espelle urina diluita; se è privato dell'acqua formerà pellet fecali secchi e il contenuto del suo retto sarà altamente iperosmotico rispetto all'emolinfa. Un'analisi dettagliata delle concentrazioni osmotiche nel complesso rettale ha permesso di suggerire un meccanismo di riassorbimento dell'acqua, presentato schematicamente in Fig. -10.7.
Si deve quindi concludere che il sistema escretore degli insetti sembra funzionare senza alcuna ultrafiltrazione iniziale, che si basa sulla secrezione primaria di potassio nei vasi malpighiani, seguita dal movimento passivo dell'acqua, e che l'acqua insieme ai soluti: Recuperato nell'intestino posteriore e nel complesso rettale: vi sono ampie prove che il movimento dell'acqua si basa sul trasporto primario di soluti.